 Jean-Pierre Raynauld, Biofísico
Jean-Pierre Raynauld, Biofísico
Université de Montréal
On peut visiter cette page en français
One can
browse this page in english
Pode-se também visualizar esta página em português
He aqui la página Web de un capítulo que aparecerá en los « Proceedings of the NATO ASI on Neurobiology », que ocurrió en Erice, Sicilia, del 2 al 12 de mayo, 1995, y que estan ahora bajo prensa donde Plenun Press: « Neurobiology: Ionic Channels, Neurons, and the Brain: editado por V.Torre y F. Ponti, 1996 ». Los que se interesan en mas detalles pueden pedir el capítulo « A Compartment Model for Vertebrate Phototransduction Predicts Sensitivity and adaptation », p.p.201-215, escribiendome a la dirección que aparece al final de esta página. Les mandaré una versión Postcriptum o versión PDF del capítulo por FTP electrónico o un tirado aparte. La versión Postscríptum y los tirados aparte ya son disponibles .
Este documento se ve mejor desde Netscape 2.0. Al escoger los íconos aparecerán las imágenes y gráficos.
Un modelo de compartimentos de la fototransducción en los vertebrados
Resumen
Un modelo de compartimentos ha sido desarrollado para explicar las diferentes propiedades de los conos y de los bastones en la sensibilidad y en la adaptación. En el caso de los conos, el compartimento natural es el pliege de la membrana plasmática; mientras que en el caso de los bastones, el compartimento es definido empíricamente. En este modelo, la cantidad de compartimentos controla la amplitud de la respuesta a la acción de un fotón; el tamaño del compartimento determina la posición de la curva « Intensidad-Respuesta » sobre el axis de las intensidades. Utilisando el teorema de ergodicidad, fue possible predecir las propiedades de la adaptación a partír de la cantidad de compartimentos y de la constante de tiempo de la fase decreciente de la respuesta a un fotón.
Introducción
 Soy un biofísico al cual le interesa saber porque los
conos y los bastones de los vertebrados presentan sensibilidades y propiedades
de adaptación diferentes, a pesar de que la bioquímica de estos fenómenos sea
tan parecida en estas dos clases de fotoreceptores. [1]
Soy un biofísico al cual le interesa saber porque los
conos y los bastones de los vertebrados presentan sensibilidades y propiedades
de adaptación diferentes, a pesar de que la bioquímica de estos fenómenos sea
tan parecida en estas dos clases de fotoreceptores. [1]
Como punto de partida, utilízo las diferencias anatómicas conocidas en la estructura externa de los fotoreceptores, magnificamente illustradas por Alfieri, R.et al. [2] (permisión requirida).
Figura 1: Bastones illustrando
el apilamento de discos en una concha cilíndrica.
 La diferencia principal es que
mientras en los conos los discos son plieges de la membrana plasmática, en los
bastones, los discos son cerrados y apilados en una concha cilíndrica.
La diferencia principal es que
mientras en los conos los discos son plieges de la membrana plasmática, en los
bastones, los discos son cerrados y apilados en una concha cilíndrica.
Ademas, un estudio tridimensional muestra que los plieges de los conos son estructuras casi completamente cerradas con una abertura de unos 20 grados en su circonferencia [3].
Figura 2: Illustración de un cono mostrando
que los plieges son parte integrante de la membrana plasmática.
Este modelo propone la existencia de compartimentos que limitan en volumen las reacciones bioquímicas consequentes de la captura de un fotón. Yo quisiera demostrar que en el caso de los conos, las membranas que forman los plieges determinan tambien el volumen o tamaño del compartimento. En el caso de los bastones, el compartimento es dicho « operacional », concepto que definiré mas lejos. Dos factores y una hipótesis son importantes para la discusión que sigue:
- El tiempo de subida de
la respuesta a un fotón.
- El coeficiente de
difusión de las moléculas implicadas en la bioquímica.
- En un compartimento dado, la acción de un fotón es máxima.
Intensidad-Respuesta
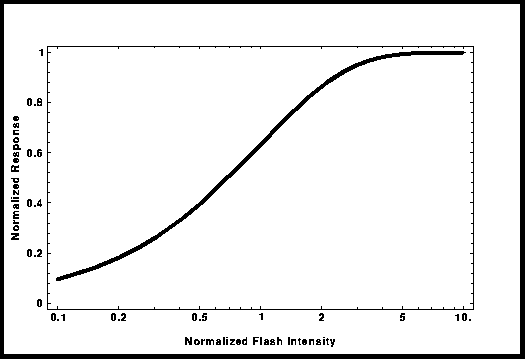 La
bioquímica responsable de la fototransducción ha sido estudiada intensamente
durante los ultimos 15 años. El resultado esta bien illustrado en el esquema de
Nakatani. La idea de que los segmentos externos de los
fotoreceptores estan compartimentados no es nueva. Desde 1981, con el propósito
de explicar la curva experimental de « Intensidad-Respuesta », Lamb et al. [4] dearrollaron el
modelo de « oclusión total ». La curva resultante, llamada de « Saturación
exponencial » puede ser utilisada en vez de la curva de « Michaelis-Menten »;
su expresión matemática es: R/Rmax = 1 - Exp[-kI], donde « I » es la intensidad
del flash y « k », un parámetro de ajuste. En este modelo, intentaré obtener la
cantidad de compartimentos y sus tamaños. De estos resultados obtendré la
respuesta a un fotón y el valor del parámetro « k ».
La
bioquímica responsable de la fototransducción ha sido estudiada intensamente
durante los ultimos 15 años. El resultado esta bien illustrado en el esquema de
Nakatani. La idea de que los segmentos externos de los
fotoreceptores estan compartimentados no es nueva. Desde 1981, con el propósito
de explicar la curva experimental de « Intensidad-Respuesta », Lamb et al. [4] dearrollaron el
modelo de « oclusión total ». La curva resultante, llamada de « Saturación
exponencial » puede ser utilisada en vez de la curva de « Michaelis-Menten »;
su expresión matemática es: R/Rmax = 1 - Exp[-kI], donde « I » es la intensidad
del flash y « k », un parámetro de ajuste. En este modelo, intentaré obtener la
cantidad de compartimentos y sus tamaños. De estos resultados obtendré la
respuesta a un fotón y el valor del parámetro « k ».
Figura 3: Curva de «
Intensidad-Respuesta »
en el modelo « oclusión total ».
Adaptación Weber-Fechner
Bajo una
illuminación cuya intensidad no es suficiente para reducir la densidad del pigmento
visual, la reducción de sensibilidad sigue una ley dicha de « Weber-Fechner »,
enunciada hace mas de un siglo. Esta ley muestra que la sensibilidad
normalizada decrece con la intensidad de la luz ambiente " I " segun
la relación « Io/(I + Io) », donde « Io » es la luz ambiente requirida para
reducir la sensibilidad de un factor un medio. Es interesante observar ese un
siglo antes de Weber y Fechner, el francés Pierre Bouguer era el primer para
medir el efecto de la luz ambiente en umbral en el estado photopic. Su medida
del umbral del incremento era 1/64 de la intensidad del fondo, un valor todavía
validado hoy. Las propiedades de adaptación de un modelo de compartimentos no
han sido estudiadas antes. Mi análisis es del tipo caja negra: despues de una
isomerización que perturbe el compartimento, las reacciones que rigen la
respuesta normalizada regresan a la situación de equilibrio que existía en la
oscuridad. Entre todas las reacciones químicas que participan en este proceso,
una sola es la limitante y hago la hipótesis que la forma de la fotocorriente
refleja esa reacción controladora de la sensibilidad. Entonces importa definir
la forma de la respuesta a un fotón y escogí la exponencial. En este esquema,
despues de una isomerización, la corriente aumenta rapidamente y decrece de
manera exponencial caracterizada por una constante de tiempo. Esto ne es una
mala aproximación puesto que varios conos y bastones presentan tales
respuestas. [5].
En mi análisis de un
systema de compartimentos, me refiero a un teorema de mecánica estadística
llamado teorema de ergodicidad que se enuncia asi : En un proceso
estochástico estacionario, un gran numero de observaciones hechas en un solo
sistema en « N » momentos arbitrarios tienen las mismas propiedades
estadísticas que las observaciones hechas sobre « N » sistemas en un momento
dado. Si me es permitida la equivalencia entre compartimento y sistema,
puedo hacer el análisis de un compartimento bajo una luz dada y predecir la
conducta del fotoreceptor, que en su totalidad, es una estructura formada de
una cierta cantidad de compartimentos.
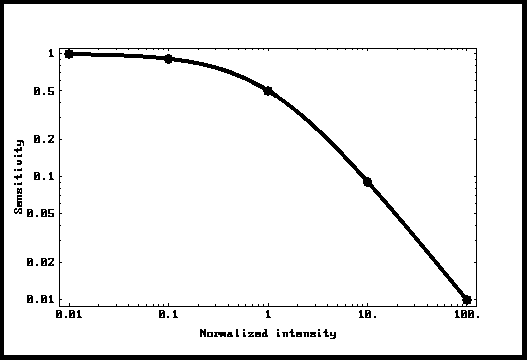
Despues de una
isomerización que produce una respuesta máxima (normalizada a 1) y decreciente
exponencialmente según una constante de tiempo « T », la amplitud de la
respuesta a un segundo fotón depende del tiempo pasado desde el fotón anterior
y sera igual a « 1 - Exp[-t/T] ». Puesto que los intervales entre las
isomerizaciones obedecen a la ley de Poisson, es posible simular una intensidad
ambiente dada y encontrar la respuesta mediana del compartimento. Esto fue
hecho en el programa de cálculo « Mathematica » y los resultados aparecen en
los puntos de la figura 4. Se puede ver que los puntos se superponen
exactamente a la curva Io/(I-Io). La conclusion es que un tal sistema se
comporta exactamente como lo predice la ley « Weber-Fechner ». Esto
representa una contribución mayor de este análisis.
Figura 4: Sensibilidad en función de la
luz ambiente,
La curva « Weber-Fechner ».
El parámetro « Io », que
hasta ahora servía unicamente para ajustar la curva a los datos experimentales,
toma ahora una significación particular ligada a la estructura del segmento
externo, es decir a la cantidad de compartimentos y a la constante de tiempo de
la respuesta a un fotón. La relación siendo Io=N/T, donde « N » es la cantidad
de compartimento y « T », la constante de tiempo de la decreciente exponencial.
El resultado final es que,
del conocimiento del tamaño del compartimento, de la cantidad de compartimento
y de la constante de tiempo asociada a la respuesta a un fotón, podemos
predecir la amplitud de la respuesta unitaria y localizar absolutamente la
posición de las curvas « Intensidad-Respuesta » y « Weber-Fechner » segun el
axis de las intensidades.
Resultados y Discusión
El caso de los conos
Por una
parte, la respuesta de los conos de mamífero a un flash de baja intensidad
apunta con una latencia de 50 msec.; por otra parte, la iodopsina, la
transducina y la fosfodiesterase, todas proteinas membranarias implicadas en la
reacción en cascada, tienen coeficientes de difusión de 0,5 a 2 micróns
cuadrados por sec.. Moviendose en un espacio bidimensional, muy pocas de estas
moléculas activadas pueden salir del pliege membranario, cuyo diámetro es de
1,5 a 2 micróns, en un lapso tan corto de tiempo. Para el GMPc, que esta en
solución y cuyo coeficiente de difusión es mas grande, hay que considerar en
que medida las moléculas situadas en discos adjacentes al disco donde se
produjo la isomerización, salen de estos y vienen a hydrolizarse en el disco
activo. Hago la hipótesis de que, falta de tiempo, esta situación es un evento
raro. En el interior del disco activo, se puede hacer un cálculo simple y
mostrar que la cantidad de fosfodiesterase activadas puede exceder la cantidad
de moléculas libres de GMPc. Existe entonces una posibilidad real de que una
sola isomerización cause la cerradura de todos los canales iónicos de un disco
o de un pliege de cono.
El primer resultado de este
análisis es que, si en los conos, el disco forma un compartimento y que la
respuesta al interior de ese compartimento es máxima, entonces la amplitud de
la respuesta a un fotón es igual a 1/N, donde « N » es la cantidad de discos.
La microscopía electrónica nos dice que la cantidad de discos por micrón es mas
o menos de 33. La cantidad total de discos es entonces igual al largo del
segmento externo multiplicado por 33. En el caso del cono de tortuga, ese
número es mas o menos de 500; la respuesta predicha teóricamente despues de la
acción de un fotón es un cambio de 0,2%, el valor obtenido experimentalmente es
de 0,16% [6] en
acuerdo con la predicción. Ayuda valiosa reciente
de 2006[7] ofertas
de los datos al modelo 1/N. En ratones genético
modificados uno puede obtener los conos que tienen segmento externo
de diversas longitudes y la sola respuesta del fotón de estos conos fue medida.
Los ratones Gt alpha -/- tiene conos S and M con un segmento
externo 13.4 micrones de largo y demuestran una sensibilidad a un
fotón de 0.2 y 0.22 % de la respuesta máxima. Según el modelo 1/N, estos
conos deben demostrar una respuesta a un fotón de % de 1/(13.4 x 33) o 0.23 %.
Los ratones Nrl -/- tiene conos S con un segmento externo
7.1 micrones de largo y una sola respuesta del fotón de 0.44% cuando el modelo
predice 1/(7.1 x 33) o 0.43%. Es difícil pedir una ayuda mejor para mi modelo.
Otro resultado de este análisis es una hipótesis
para explicar porque los conos son, por lo general, cortos. Effectivamente, si
el hecho de agregar un disco aumenta la probabilidad de captura de un fotón, la
respuesta disminuye de un factor 1/N a 1/(N+1). La naturaleza tuvo que hacer un
compromiso entre capturar mas fotones y dar una respuesta adequada a la captura
de esos fotones.
Conociendo las dimensiones
del compartimento, es decir del disco o pliege plasmático, es posible calcular
el valor del parámetro « k » en la fórmula de saturación exponencial [8]. Del valor de la
densidad óptica del segmento externo y del de la eficacia quántica, se
demuestra que « k » es igual a 0,00059 veces el cuadrado del diámetro del
segmento externo expresado en micrones. Cuando derivamos esta fórmula,
consideramos que el cono estaba estimulado transversalmente, lo que coresponde
a la situación experimental. Esa derivación es válida tambien en el caso de un
estímulo axial corespondiente a la situación fisiológica mientras el segmento
externo no sea demasiado largo. En el caso de los conos de Macaca, el diámetro
es mas o menos 1,5 micrón, el valor de « k » calculado es 0,001 micrones
cuadrados, mientras que el valor obtenido experimentalmente es de 0.0004 [9] osea mas o menos la
mitad.
Una estimación de la
intensidad (medida en isomerización por sec.) necesaria para reducir la
sensibilidad de 50% es obtenida dividiendo la cantidad de compartimentos (800
en los conos de tortuga) por la constante de tiempo que caracteriza la
decreciente de la respuesta a un flash de baja intensidad. El cálculo da mas o
menos 7300 para el valor de « Io ». Experimentalmente se obtienen valores entre
2000 y 16000 por sec. [10].
El caso de los bastones
En el caso
de los bastones, el compartimento no esta definido por membranas como en el
caso de los conos sino que por la combinación del hecho de que las proteinas
activadas en la fototransducción tienen todas un tiempo de vida limitado y que
el GMPc que controla la abertura de los canales tiene un coeficiente de
difusión reducido en los alrededores del segmento externo. La respuesta de un
bastón de mamífero apunta todos los 100 msec. mas o menos, esto permite la
activación de mas moléculas de transducinas y de fosfodiesterase. La acción de
la bioquímica puede dejar el espacio inter-discos y extenderse longitudinalmente.
La geometría de apilamiento de los discos en el segmento externo de los
bastones es importante. Para propagarse longitudinalmente, la reducción de GMPc
debe reducir la concentración entre cada disco a lo largo de su recorrido. Esto
limita la acción de la bioquímica. Estamos en presensia de un comportamiento
dicho « operacional ».
Cálculos complejos en
dinámica de los fluidos serian requiridos para determinar la concentración
longitudinal de GMPc en los alrededores de la membrana plasmática despues de
una isomerización en un disco. Sin embargo, una formula empírica fue obtenida
del análisis de la respuesta a un fotón en varias especies. Esta relación dice
que la superficie de membrana plasmática donde los canales iónicos estan
cerrados es igual a la de un disco (o capas) del segmento externo. Un cálculo
simple dice que al largo implicado debe ser igual al diámetro dividido por dos.
Para los bastones de sapo, de rata, de monos y tambien de humano, la reducción
de corriente predicha es 4 a 5%, osea el valor obtenido experimentalmente.
Cálculos similares a los de los conos dan un valor de « k » para los bastones
igual a 0,01 veces el cubo del diámetro del segmento externo expresado en
micrones. Este cálculo permite localizar la curva « Intensidad-Respuesta » de
manera absoluta sobre el axis de las intensidades. Para los bastones de
mamífero, el valor calculado es de 0,08 micrones cuadrados; los valores
experimentales varían entre 0.006 y 0.07. [8,11]
La cantidad de compartimentos
" N " de un bastón es obtenido al dividir el largo total del segmento
externo por d/2, esto da un valor cerca de 20. « Io » en la relación «
Weber-Fechner » es entonces 20 dividido por la constante de tiempo de la
decreciente exponencial. Para los bastones de los vertebrados superiores
(hombres, monos), la predicción da un « Io » entre 110 y 113 isomerizaciones
por sec. mientras que los resultados experimentales dan un valor entre 100 y
120 isomerizaciones por sec. [7,10]
Conclusión
El análisis
de la estructura geométrica del segmento externo de los fotoreceptores de
vertebrados a dado resultados interesantes.
- En primer lugar, fue posible predecir la
respuesta a un fotón y cálcular el valor del parámetro « k » en la fórmula
de « Saturación exponencial », permitiendo la localización de la curva en
el axis de las intensidades.
- Despues, gracias al teorema de ergodicidad
fue posible predecir las propiedades de adaptación de un sistema de
compartimentos. Si la respuesta es máxima y la decreciente exponencial, el
resultado es la ley « Weber-Fechner ». Esta curva tambien esta localizada
de manera absoluta en el axis de las intensidades.
Ademas, la validez del
modelo puede ser testeada. Efectivamente, en los vertebrados a sangre fria, la
cinetica de la respuesta a un flash varía con la temperatura
Vuestros comentarios son
apreciados-correo electrónico:jean-pierre.raynauld@umontreal.ca
1. Lamb
TD, Pugh EN (1992) G-protein cascades: gain and kinetics . Trends Neurosci 15:
291-298.
2. Alfieri
R, Sole P, Gentou C, Kantelip B, Kantelip J-P . (1984) Les cellules
rétiniennes. Ed. D.G.P.L. Paris.
3.
Eckmiller MS (1987) Cone Outer Segment Morphogenesis: Taper Change and Distal
Invaginations. J Cell Biol 105: 2267-2277.
4. Lamb
TD, McNaughton PA, Yau KW (1981) Spatial spread of activation and background
desensitization in toad rod outer segments. J Physiol 319: 463-496.
5. Corson
DW, Cornwall MC, MacNichol EF, Tsang S, Derguini F, Crouch RK,Nakanishi K
(1994) Relief of opsin desensitization and prolonged excitation of rod
photoreceptors by 9-desmethylretinal. Proc Natl Acad Sci USA 91: 6958-6962.
6. Baylor
DA, Hodgkin AL (1973) Detection and resolution of visual stimuli by turtle
photoreceptors. J Physiol 234: 163-198
7 Nikonov SS,
Kholodenko R, Lem J, and Pugh EN. (2006). Physiological features of the S- and
M-cone Photoreceptors of wild-type mice from single-cell recordings.
J.Gen. Physiol. 127(4), 359-374
8. Baylor
DA., Nunn BJ, Schnapf JL (1984) The photocurrent noise and spectral
sensitivities of rods of the monkey Macaca fascicularis. J Physiol 357: 575-607
9. Schnapf
JL, Nunn BJ, Meister M, Baylor DA (1990) Visual transduction in cones of the
monkey Macaca fascicularis. J Physiol 427: 681-713 10. Baylor
DA, Hodgkin AL (1974) Change in time scale and sensitivity in turtle
photoreceptors. J Physiol 242: 729-758. 11. Kraft
TW, Schneeweis DM, Schnapf JL (1993) Visual Transduction in human
photoreceptors. J Physiol 464: 747-765 12. Lamb
TD (1984) Effects of temperature changes on toad rod photo currents. J Phsysiol
346: 557-578 Salido de mi bocal de pecesillos rojos
Burkhardt DA (1994) Light adaptation and photopigment bleaching in cone
photoreceptors in situ in the retina of the turtle. J Neurosci 14(3),
1091-1105.
Jean-Pierre Raynauld
Traducido al castellano por Mario Villalón, Biofísico.
(last updated May 24th, 2007)